It’s hard to believe we started this blog over eight years ago – all the way back when we were grad students. What a long way we’ve come. Patryk is now Director of R&D at Brain Corporation, while Michael is an assistant professor at Rutgers University. Today we are relaunching Neurevolution, with a new design and …
Category Archives: General Neuroscience
The brain’s network switching stations for adaptive behavior
I’m excited to announce that my latest scientific publication – “Multi-task connectivity reveals flexible hubs for adaptive task control” – was just published in Nature Neuroscience. The paper reports on a project I (along with my co-authors) have been working on for over a year. The goal was to use network science to better understand how …
Continue reading “The brain’s network switching stations for adaptive behavior”
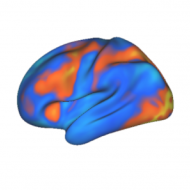
Having more global brain connectivity with some regions enhances intelligence
A new study – titled “Global Connectivity of Prefrontal Cortex Predicts Cognitive Control and Intelligence” – was published just last week. In it, my co-authors and I describe our research showing that connectivity with a particular part of the prefrontal cortex can predict how intelligent someone is. We measured intelligence using “fluid intelligence” tests, which measure …
Continue reading “Having more global brain connectivity with some regions enhances intelligence”
The evolutionary importance of rapid instructed task learning (RITL)
We are rarely alone when learning something for the first time. We are social creatures, and whether it’s a new technology or an ancient tradition, we typically benefit from instruction when learning new tasks. This form of learning–in which a task is rapidly (within seconds) learned from instruction–can be referred to as rapid instructed task …
Continue reading “The evolutionary importance of rapid instructed task learning (RITL)”
Finding the most important brain regions
When you type a search into Google it figures out the most important websites based in part on how many links each has from other websites. Taking up precious website space with a link is costly, making each additional link to a page a good indicator of importance. We thought the same logic might apply …
Cingulate Cortex and the Evolution of Human Uniqueness
Figuring out how the brain decides between two options is difficult. This is especially true for the human brain, whose activity is typically accessible only via the small and occasionally distorted window provided by new imaging technologies (such as functional MRI (fMRI)). In contrast, it is typically more accurate to observe monkey brains since the …
Continue reading “Cingulate Cortex and the Evolution of Human Uniqueness”
A Meta-Meta-Analysis of Brain Functions
Thousands of brain imaging studies are published each year. A subset of these studies are replications, or slight variations, of previous studies. Attempting to come to a solid conclusion based on the complex brain activity patterns reported by all these replications can be daunting. Meta-analysis is one tool that has been used to make sense …
Grand Challenges of Neuroscience: Day 6
Topic 6: Causal Understanding Causal understanding is an important part of human cognition. How do we understand that a particular event or force has caused another event? How do realize that inserting coins into a soda machine results in a cool beverage appearing below? And ultimately, how do we understand people’s reactions to events? The …
Joaquin Fuster on Cortical Dynamics
I recently watched this talk (below) by Joaquin Fuster. His theories provide a good integration of cortical functions and distributed processing in working and long-term memory. He also has some cool videos of likely network interactions across cortex (in real time) in his talk. Here is a diagram of Dr. Fuster’s view of cortical hierarchies: …
Combining Simple Recurrent Networks and Eye-Movements to study Language Processing
Modern technologies allow eye movements to be used as a tool for studying language processing during tasks such as natural reading. Saccadic eye movements during reading turn out to be highly sensitive to a number of linguistic variables. A number of computational models of eye movement control have been developed to explain how these variables …
Magnetoencephalography
In the dark confines behind our eyes lies flesh full of mysterious patterns, constituting our hopes, desires, knowledge, and everything else fundamental to who we are. Since at least the time of Hippocrates we have wondered about the nature of this flesh and its functions. Finally, after thousands of years of wondering we are now …
Two Universes, Same Structure
This image is not of a neuron. This image is of the other universe; the one outside our heads. It depicts the “evolution of the matter distribution in a cubic region of the Universe over 2 billion light-years”, as computed by the Millennium Simulation. (Click the image above for a better view.) The next image, …
Grand Challenges of Neuroscience: Day 2
Topic 2: Conflict and Cooperation Generally, cognitive neuroscience aims to explain how mental processes such as believing, knowing, and inferring arise in the brain and affect behavior. Two behaviors that have important effects on the survival of humans are cooperation and conflict. According to the NSF committee convened last year, conflict and cooperation is an …
Grand Challenges of Neuroscience: Day 1
Following up on MC's posts about the significant insights in the history of neuroscience, I'll now take Neurevolution for a short jaunt into neuroscience's potential future. In light of recent advances in technologies and methodologies applicable to neuroscience research, the National Science Foundation last summer released a document on the "Grand Challenges of Neuroscience". These …
History’s Top Insights Into Brain Computation
This post is the culmination of a month-long chronicling of the major brain computation insights of all time. Some important insights were certainly left out, so feel free to add comments with your favorites. Below you will find all 26 insights listed with links to their entries. At the end is the summary of the …
Continue reading “History’s Top Insights Into Brain Computation”