Humans have come to dominate the world largely due to our intelligence relative to other species. Where does this tremendous cognitive flexibility come from?
Many have considered prefrontal cortex as the origin of this boost in intelligence, yet it has remained unclear how this chunk of brain tissue could drive intelligent behavior. In a recently published study we used fMRI (functional magnetic resonance imaging) along with advanced machine learning approaches to read out what information is processed in prefrontal cortex during intelligent behavior.
We focused on one of the most remarkable abilities indicative of human intelligence: rapid instructed task learning (RITL). This is the ability to quickly learn new tasks from instructions. For instance, we use it to learn to use new technology (e.g., a new smartphone) and when learning to play a new game (e.g., Monopoly).
This ability may seem mundane because it’s familiar, but it’s actually quite remarkable both computationally and in terms of evolution. Computationally it’s remarkable because even modern computers can’t match the RITL abilities of human children. Evolutionarily it’s remarkable because other primates take much longer to learn many tasks that we can perform immediately upon hearing instructions.
We have investigated RITL in the past. What’s different here is that we were able to tie what information is in prefrontal cortex (instead of just how active it is) to how accurate a person is on a given trial. In other words, we were able to see the information processing in prefrontal cortex that drives successfully intelligent task performance .
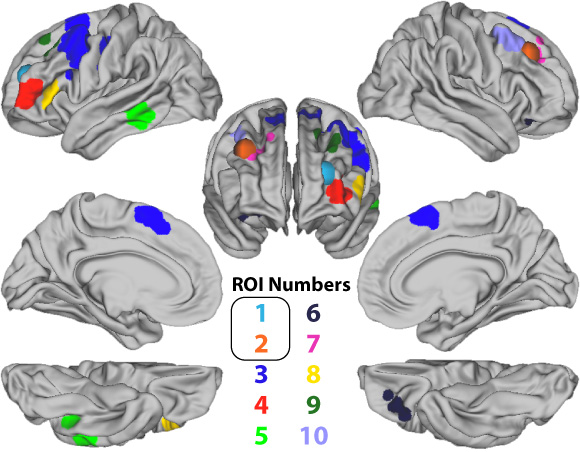
We started out finding the brain regions (regions of interest; ROI) that were more responsive to novel that practice tasks. This helped us isolate RITL from general task processing. We found a variety of prefrontal areas that were preferentially involved in novel relative to practiced task performance (Figure 1).
We wanted to test whether any of these brain regions were especially important for RITL behavior. We therefore tested if there were any regions that also 1) represented task information and 2) were sensitive to task accuracy. We found three brain regions that showed all of these properties. Surprisingly, all three regions were in left anterior prefrontal canadian roulette cortex (regions 1, 4, & 8 in Figure 1).
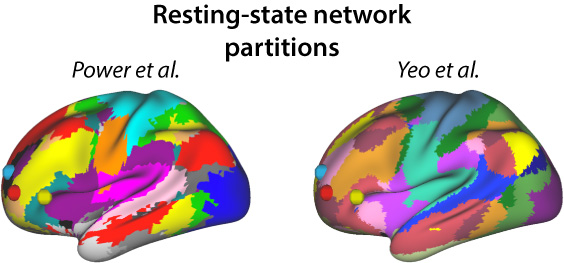
Given how close these regions were to each other, we thought they might actually be the same large brain region. When we checked their overlap with brain connectivity patterns, however, we saw that they are each part of a different network (Figure 2).
This suggests there are three nearby regions in left anterior prefrontal cortex that play an especially important role in flexible and intelligent behavior in humans.
See the full scientific article for more info.
-MWCole





 When you type a search into Google it figures out the most important websites based in part on how many links each has from other websites. Taking up precious website space with a link is costly, making each additional link to a page a good indicator of importance.
When you type a search into Google it figures out the most important websites based in part on how many links each has from other websites. Taking up precious website space with a link is costly, making each additional link to a page a good indicator of importance.

")
")
 Causal understanding is an important part of human cognition. How do we understand that a particular event or force has caused another event? How do realize that inserting coins into a soda machine results in a cool beverage appearing below? And ultimately, how do we understand people’s reactions to events?
Causal understanding is an important part of human cognition. How do we understand that a particular event or force has caused another event? How do realize that inserting coins into a soda machine results in a cool beverage appearing below? And ultimately, how do we understand people’s reactions to events?
")
")
