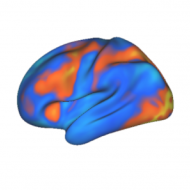
The frontoparietal control system is to the mind what the immune system is to the body. It may oversimplify the situation, but we’re finding it’s a useful metaphor nonetheless. Indeed, we’ve just published a new theory paper explaining that there is already an avalanche of evidence supporting this metaphor. Even though much work is left …
Continue reading “Frontoparietal cortex: The immune system of the mind”